DIAGNOSIS
Lung adenocarcinoma with MET exon 14 skipping
Follow-up:
Upon clinical staging, the patient was found to be stage IIB (cT3 N0 M0), for which the recommendation is local control followed by adjuvant immunotherapy. After thoracic surgery evaluation, he was determined to not be a surgical candidate and was subsequently treated with chemotherapy (carboplatin, paclitaxel) and radiation therapy. Immunotherapy will be considered depending on the response to chemotherapy and radiation therapy. Targeted therapy for the MET exon 14 skipping with capmatinib remains a possibility if metastatic disease develops in the future.
DISCUSSION
Overview of molecular alterations in lung adenocarcinoma [1-7]:
Adenocarcinoma is the most common subtype of non-small cell lung carcinoma (NSCLC). See Table 1 for a list of the most common molecular driver alterations in lung adenocarcinoma. A significant proportion of tumors also have alterations in a variety of tumor suppressor genes, such as TP53 (45-50%) and STK11 (15-20%). Many of these molecular alterations have targeted therapies available. For some alterations, secondary resistance alterations (mutations, amplifications, activation of alternate pathways, etc.) may arise with targeted therapy. This phenomenon is most commonly reported with EGFR (ex. T790M mutation) and ALK (ex. G1269A, G1202R mutations) alterations. KRAS mutations are more common in older patients who are heavy smokers, while many of the other alterations are more common in younger patients, non-smokers or light smokers, and patients of Asian descent.
From a therapeutic perspective, KRAS mutations result in resistance to anti-EGFR therapies. While no FDA approved therapies are currently available, MEK inhibitors are currently under investigation. BRAF mutations, particularly V600E, are uncommon but may respond to BRAF and/or MEK inhibitors. A variety of EGFR mutations have been reported, which can be broadly divided into tyrosine kinase inhibitor (TKI) sensitive or resistant, based on their response to earlier generation TKIs (ex. gefitinib, erlotinib). The most common EGFR mutations, L858R and in-frame exon 19 deletions, are considered TKI sensitive. More common TKI resistant mutations include T790M and in-frame exon 20 insertions, which may respond to later generation TKIs (ex. afatinib, osimertinib). Multiple fusions have been reported in lung adenocarcinoma, including ALK (most commonly EML4/ALK), ROS1 (most commonly ROS1/CD74), RET, NTRK, and NRG1 (most commonly NRG1/CD74), all of which have potential targeted therapies, several of which are FDA approved. Therapy for MET alterations will be discussed below. Aside from KRAS and BRAF, many of these altered genes function as receptor tyrosine kinases (RTK), and therefore may respond to non-specific or gene specific TKIs. Most FDA approved treatments are approved specifically for metastatic and/or resistant/refractory disease, with some available as first-line therapy in these settings.
MET exon 14 skipping in non-small cell lung carcinoma (NSCLC) [8-16]:
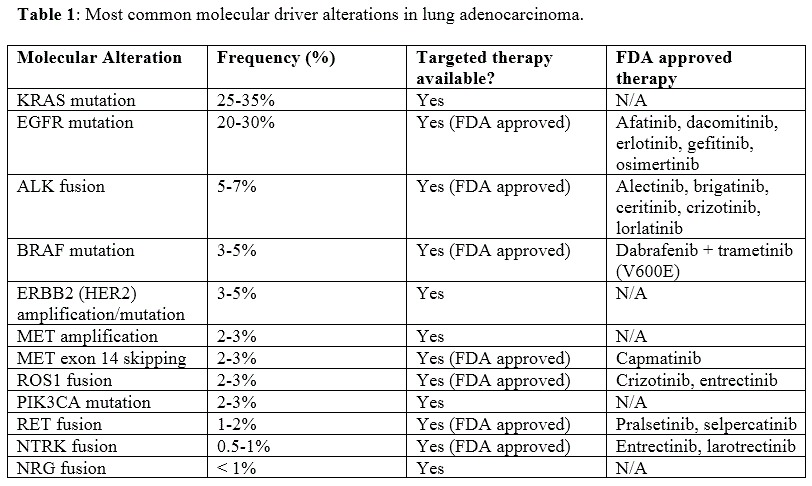
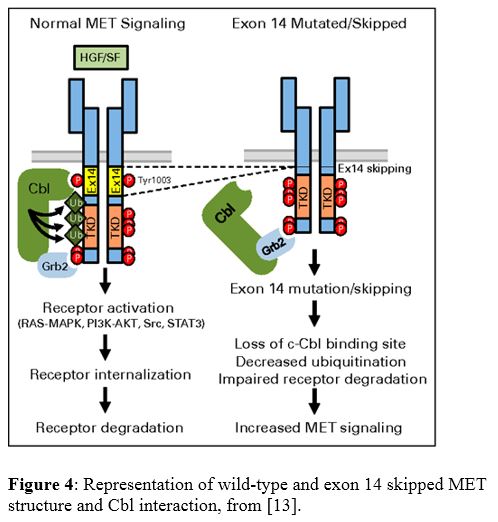
MET, also known as hepatocyte growth factor receptor (HGFR), is an oncogene receptor tyrosine kinase (RTK) which activates a variety of pathways, including MAPK, PI3K/AKT, SRC, and STAT. MET exon 14 skipping is uncommon, but has been reported in up to 3% of NSCLC, mostly in adenocarcinoma and sarcomatoid carcinoma. This alteration has also been reported to be more common in older female non-smokers. There is no data regarding the prognostic impact of this alteration. As an RTK, tumors with this alteration have shown response to crizotinib, other MET-specific tyrosine kinase inhibitors (TKI), and a recently FDA approved drug specifically for NSCLC with this alteration, capmatinib. Mechanistically, exon 14 of MET contains a juxtamembrane domain with a binding site for Cbl, which is an E3 ubiquitin-ligase. In the wild-type state, Cbl binds to residues in exon 14, which allows for ubiquitination of MET and subsequent receptor internalization and degradation. Loss of exon 14 prevents this action, resulting in impaired degradation and increased MET signaling and activation of multiple pathways (Figure 4). The most commonly associated causative alteration is single nucleotide variants (SNV) of the splice donor site, followed by SNVs of the splice acceptor site and insertions/deletions of the splice donor site. These mutations impair the normal splicing process around exon 14, resulting in its skipping and exclusion from the mRNA and protein (Figure 5).
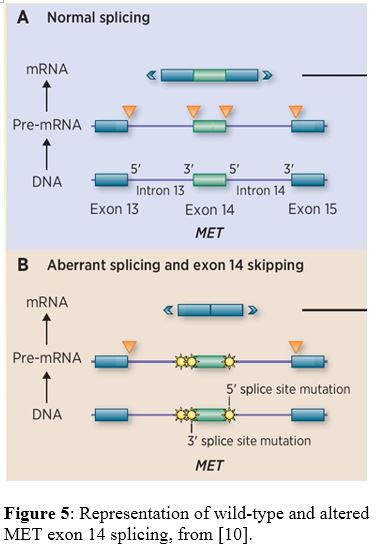
Overview of mRNA splicing [17-18]:
The spliceosome complex includes multiple small nuclear ribonucleoproteins (snRNPs) and is critical for the proper processing of RNA. Genes are made up of exons and introns, of which the exons are the defined coding regions for the target gene protein. RNA splicing removes the introns and forms mature mRNA, which is then able to form a functional protein. Without functional splicing, proteins may become over- or under-stabilized by gaining or losing material.
Critical sequences within the intron allows for proper binding of the spliceosome snRNPs (U1, U2, etc.) and subsequent functional splicing. These sequences include the 5' donor site (GU, n+1/2 positions), 3' acceptor site (AG, n-1/2 positions), and branch-point site (A within somewhat variable sequence, upstream from 3' AG). The 5' donor site is recognized by U1, and the branch-point site is recognized by U2. A complex of U4, U5, and U6 replace U1 and bring the 5' exon in proximity to the branch-point site. The 5' -OH group, facilitated by U5, attacks the branch-point site A to release the 5' exon and form a lariat-type structure. The 5' exon is brought in proximity to the 3' exon, and U5 again facilities a 5' attack to join the two exons and release the intron. See Figure 6 for a pictorial representation. Of note, there is also some level of conservation in the nucleotides adjacent to the consensus splice sites, both exonic and intronic. The spliceosome components are formed from combinations of protein coding genes and ribosomal RNA (ex. U2 - SF3A1, SF3B1, 12S rRNA).
REFERENCES
![]() Contributed by John M. Skaugen, MD, Tim D. Oury, MD, PhD, Yuri E. Nikiforov, MD, PhD
Contributed by John M. Skaugen, MD, Tim D. Oury, MD, PhD, Yuri E. Nikiforov, MD, PhD